溶菌酶测定实验报告(多糖寡糖蛋白酶在促生长、助消化提升免疫力方面重要性您知道吗?)
Posted
篇首语:再冷的石头,坐上三年也会暖。本文由小常识网(cha138.com)小编为大家整理,主要介绍了溶菌酶测定实验报告(多糖寡糖蛋白酶在促生长、助消化提升免疫力方面重要性您知道吗?)相关的知识,希望对你有一定的参考价值。
溶菌酶测定实验报告(多糖寡糖蛋白酶在促生长、助消化提升免疫力方面重要性您知道吗?)

谭崇桂1,冷向军*1,2,李小勤1,2,粟雄高1,刘波1,柴仙琦1(1上海海洋大学水产与生命学院,上海 201306,中国)(2农业部淡水水产种质资源重点实验室,上海,201306,中国)
摘要:为考察饲料中添加β-葡聚糖、黄芪多糖、甘露寡糖和蛋白酶对凡纳滨对虾生长性能、血清非特异性免疫及消化酶活性的影响,在基础饲料(对照组)中添加0.2%β-葡聚糖、0.05%黄芪多糖、0.4%甘露寡糖、175mg/kg蛋白酶PT,选取初始体重为4.55±0.08g的凡纳滨对虾(Litopnnaus vannamei)1000尾,分为5处理组,每处理组4重复,每重复50尾虾,分别饲喂上述5组饲料6周。
结果表明:与对照组相比,饲料中添加0.2%β-葡聚糖、0.4%甘露寡糖及175mg/kg蛋白酶PT可分别提高凡纳滨对虾增重率16.19%、12.78%、11.25%(P<0.05),提高蛋白质效率8.67%、8.32%、8.04%(P<0.05),降低饲料系数0.10、0.09、0.09(P<0.05)。
各处理组在肌肉水分、灰分、粗脂肪和粗蛋白含量上无显著差异(P>0.05)。
在血清非特异性免疫方面,各多糖、寡糖和蛋白酶添加组均显著提高血清酚氧化酶活力(P<0.05),但在丙二醛含量方面无显著差异(P>0.05);
添加0.4%甘露寡糖可提高溶菌酶、碱性磷酸酶活力(P<0.05);添加0.2%β-葡聚糖、0.4%甘露寡糖可显著提高超氧化物歧化酶活力(P<0.05)。
在消化酶活性方面,添加0.05%黄芪多糖及175mg/kg蛋白酶PT可显著提高肝胰脏蛋白酶活性(P<0.05),添加175mg/kg蛋白酶PT显著提高肠蛋白酶活性(P<0.05),添加0.2%β-葡聚糖及0.4%甘露寡糖显著提高胃蛋白酶活性(P<0.05),添加0.2%β-葡聚糖、0.4%甘露寡糖及175mg/kg蛋白酶PT显著提高肝胰脏脂肪酶活性(P<0.05)。
上述结果表明,饲料中添加0.2%β-葡聚糖、0.4%甘露寡糖可提高凡纳滨对虾生长性能、血清非特异性免疫,改善消化酶活性;添加175mg/kg蛋白酶PT可提高凡纳滨对虾增重率,改善消化酶活性;添加0.05%黄芪多糖对凡纳滨对虾增重率、血清非特异免疫无显著影响。
近年来水产养殖业快速发展的同时,也出现了环境污染,水产动物抗病力下降和病害加剧等问题;
同时由于鱼粉价格上涨,植物蛋白的大量使用,导致饲料消化利用率降低。寻求绿色、环保、高效的添加剂已成为水产动物营养与饲料学研究的热点。
在这方面,免疫多糖、功能性寡糖和酶制剂因具有调节肠道微生物菌群,提高机体免疫功能,提高营养物质消化率的作用而受到重视。
β-葡聚糖作为免疫增强剂在水产动物上已有较多报道[1-2]。
在虾类方面,饲料中添加0.2%β-葡聚糖可显著提高凡纳滨对虾增重率[3],提高溶菌酶、超氧化物歧化酶和酚氧化酶活性[4];
添加10g/kg的β-1,3-葡聚糖,可以提高斑节对虾(Penaeus monodon)感染白斑综合症病毒后的存活率[5];
黄芪多糖[6-7]、甘露寡糖[8-10] 、蛋白酶[11-16]在鱼类的研究中也有一些报道,但在虾类中均研究报道较少。
本试验以凡纳滨对虾为研究对象,在饲料中分别添加0.2%β-葡聚糖、0.05%黄芪多糖、0.4%甘露寡糖、175mg/kg蛋白酶PT,考察并比较几种添加剂对凡纳滨对虾生长性能、肌肉成分、消化酶活性、血清非特异性免疫指标影响,为多糖、寡糖、蛋白酶在对虾饲料中的合理应用提供依据。
1 材料与方法
1.1 试验设计与饲料
在基础饲料中分别添加0.0%(对照组)、0.2%β-葡聚糖(由广州信豚生物科技有限公司提供,β-葡聚糖有效含量25%)、0.4%甘露寡糖(由北京奥特奇生物科技有限公司提供,甘露寡糖含量为12%)、 175mg/kg蛋白酶PT(由加拿大JEFO公司提供,为一种耐高温的碱性蛋白酶)、0.05%黄芪多糖(上海朝翔生物技术有限公司提供,主要成分为35%黄芪多糖及11%黄芪甲甙),共5个处理组。
基础饲料组成及营养成分见表1。饲料原料均过60目筛,采用逐级混匀的方法将原料混合,用绞肉机制成直径为1mm的硬颗粒沉性饲料,95℃后熟化25min,自然风干,4℃保存备用。
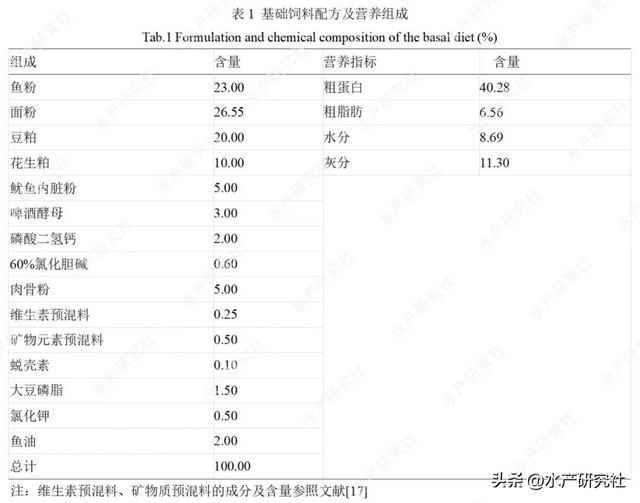
1.2 试验用虾及饲养管理
试验用虾购于上海市青浦区水产养殖场。对虾运回后,暂养在网箱(2.5m×1.2m×1.0m)中,网箱置于水泥池(5.0m×3.0m×1.2m)中,用基础饲料驯化一周,以适应饲料与环境。
选用均重为4.55g左右、规格均匀、体质健壮的1000尾虾开始正式试验。试验虾共分为5组,每组四重复,每个网箱为一个重复,每重复50尾虾。
每天投喂三次(5:30、12:30、18:30),日投饵量约占虾体重的3%~5%,早晚投喂量约占总投喂量的70%,并根据实际情况做适当调整。
每隔3d吸污与换水,保持水质。养殖期间水温25~30℃,DO>5mg.L-1,PH:7~8,NH4+-N<0.2mg/L,养殖时间为2011年8月6日至2011年9月16日,共6周,合计42d。养殖试验在上海海洋大学南汇特种水产养殖场进行。
1.3 生长性能
养殖试验结束后,试验虾饥饿24h,称重,计算增重率(RWG)、蛋白质效率(PER)、成活率(RS)、饲料系数(FCR)。
RWG=100×(W1-W0)/W0
PER=(W1-W0)/WP
RS=100×NS/NT
FCR=FT/(W1-W0)
式中:W0为平均初始体重(g);W1为平均终末体重(g);WP为蛋白质摄取量(g);FT为总投饲量(g);NS为成活尾数;NT为总尾数。
1.4 肌肉常规成分
养殖结束后,每重复采虾5尾,取肌肉样品-20℃冷冻保存用于常规成分分析,样品分析采用国际标准方法[18]。
水分测定采用105℃烘干恒重法,粗蛋白测定采用凯氏定氮法,粗脂肪测定采用氯仿-甲醇法,粗灰分测定采用550℃灼烧法。
1.5 血清非特异性免疫指标
养殖试验结束后,试验虾饥饿24h,每池随机取虾15尾,自虾的头胸甲后插入围心腔取血淋巴液,置于离心管中,4℃,5000r/min,离心10min,取上清液用作血清非特异性免疫指标的测定。
其中溶菌酶、超氧化物歧化酶、碱性磷酸酶、丙二醛均采用南京建成生物工程研究所生产的试剂盒测定。
溶菌酶:采用比浊法,溶菌酶含量(U/mL)=(测定管透光度UT2-测定管透光度UT1)/(标准管透光度ST2-标准管透光度ST1)×标准管浓度(200U/mL)×样本测试前稀释倍数;
超氧化物歧化酶(SOD):采用黄嘌呤氧化酶法,1个SOD活力单位(U)定义为每mL血清中SOD抑制率达50%时所对应的SOD量;
碱性磷酸酶(AKP):1个单位(U)定义为每mL血清在37℃与基质作用15分钟产生1mg酚;
丙二醛(MDA):采用硫代巴比妥酸(TBA)法;酚氧化酶:以左旋多巴为底物,参照Ashida[19]的方法进行操作。
将3mL的磷酸盐缓冲液(0.1mol/L,pH=6.4)、100μL血清及100μL左旋多巴在室温下混匀,490nm波长下,每2min读取光度密值。
以OD490对反应时间作图,以实验条件下每分钟光密度值增加0.001为1个活性单位(U)。
1.6消化酶
在解剖盘的冰浴中解剖虾,取胃、肠道、肝胰脏等组织,-20℃保存。测定时4℃解冻,,加入10倍体积的预冷重蒸水,4℃匀浆,离心10min(5000r/min),取上清液,即为粗酶液。
蛋白酶参照DIVAKARAN[20]方法进行测定。胃、肠、肝胰脏蛋白酶的活性为1mg鲜组织在pH分别为3.0、7.2、9.8时每分钟水解酪蛋白产生1μg酪氨酸为一个酶活性单位(U);
脂肪酶、淀粉酶测定均采用南京建成生物工程研究所生产的试剂盒,脂肪酶单位定义为:在37℃条件下,每g组织蛋白在本反应体系中与底物反应1min,每消耗1μmol底物为一个酶活力单位(U/g);
淀粉酶的测定采用碘-淀粉比色法,每g组织蛋白在37℃与底物作用30min,水解10mg淀粉定义为1个淀粉酶活力单位(U/g)。
1.7 数据统计
试验结果数据以平均值±标准差表示,采用SPSS16.0进行差异显著性分析和多重比较(Ducan氏法),差异显著水平为P<0.05。
2 结果
2.1对生长性能及饲料利用的影响
多糖、寡糖、蛋白酶对凡纳滨对虾生长性能及饲料利用的影响见表2。
与对照组相比,饲料中添加0.2%β-葡聚糖、0.4%甘露寡糖、175mg/kg蛋白酶PT分别提高凡纳滨对虾增重率16.19%、12.78%、11.25%(P<0.05),提高蛋白质效率8.67%、8.32%、8.04%,降低饲料系数0.10、0.09、0.09(P<0.05);黄芪多糖也有提高增重率及蛋白质效率,降低饲料系数的趋势,但与对照组差异不显著(P>0.05)。
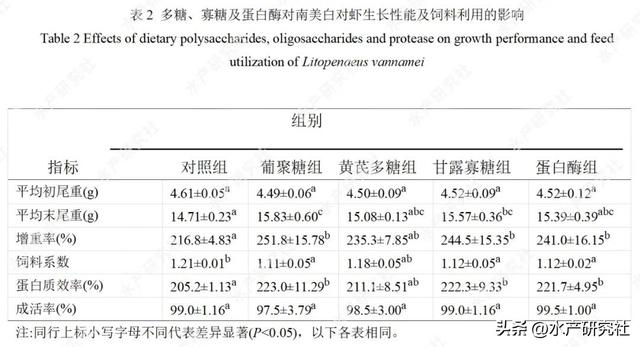
2.2对肌肉成分的影响
多糖、寡糖、蛋白酶对凡纳滨对虾肌肉成分的影响见表3。各组间水分、粗蛋白、粗脂肪、粗灰分无显著差异(P>0.05)。
2.3对血清免疫指标的影响
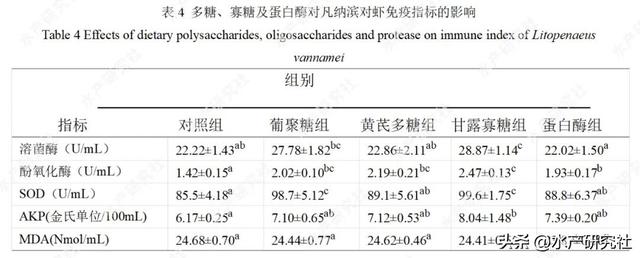
多糖、寡糖、蛋白酶对凡纳滨对虾血清免疫指标的影响见表4。
与对照组相比,饲料中添加0.4%甘露寡糖可显著提高溶菌酶及碱性磷酸酶活性(P<0.05);
添加0.2%β-葡聚糖、0.4%甘露寡糖可显著提高超氧化物歧化酶活性(P<0.05);各添加剂组均显著提高酚氧化酶活性(P<0.05),但在丙二醛含量方面无显著差异(P>0.05)。
2.4 对消化酶活性的影响
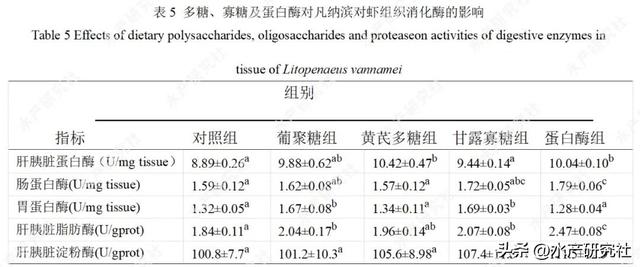
由表5可知,饲料中添加0.05%黄芪多糖、175mg/kg蛋白酶PT可显著提高凡纳滨对虾肝胰脏蛋白酶活性(P<0.05);添加175mg/kg蛋白酶PT可显著提高肠蛋白酶活性(P<0.05);
添加0.2%β-葡聚糖、0.4%甘露寡糖可显著提高胃蛋白酶活性(P<0.05);添加0.2%β-葡聚糖、0.4%甘露寡糖、175mg/kg蛋白酶PT显著提高肝胰脏脂肪酶活性(P<0.05);各组在肝胰脏淀粉酶活性方面无显著差异(P>0.05)。
3 讨论
3.1 β-葡聚糖对凡纳滨对虾生长和血清非特异性免疫的影响
β-葡聚糖作为免疫增强剂在捉进水产动物生长、增强免疫力方面已有较多研究报道[1,21]。
在对虾的研究中表明,在饲料中分别添加0.2%[3]及0.4%β-葡聚糖[22],均可显著提高凡纳滨对虾增重率。
本试验也获得相似效果,饲料中添加0.2%β-葡聚糖可提高凡纳滨对虾增重率16.19%,降低饲料系数0.10(P<0.05)。
有关β-葡聚糖促生长作用机制尚不清楚,推测β-葡聚糖可被消化提供能量,从而起到节约蛋白的作用[23]。
对虾免疫系统中存在着识别β-葡聚糖的结合蛋白,当两者相结合时,结合物可与颗粒细胞膜的受体相结合,从而激活免疫系统[24]。
研究表明,间断性投喂β-葡聚糖可以使凡纳滨对虾的免疫力维持在一个较高的水平,消除免疫疲劳[25];
在饲料中添加2g/kg酵母细胞壁及2g/kg β-葡聚糖可提高凡纳滨对虾溶菌酶、超氧化物歧化酶和酚氧化酶活性,提高机体抗病能力[4];
添加0.09%β-1,3-葡聚糖,可显著提高大黄鱼血清溶菌酶活力,头肾白细胞的吞噬活性,显著提高大黄鱼被哈维氏菌感染后的成活率(P<0.05)[1];
用0.5mg/ml、1mg/ml β-葡聚糖悬浮夜浸泡斑节对虾幼虾,可提高酚氧化酶活力及抗菌活力[26]。
Chang等[5]研究表明:摄食10g/kg的β-1,3-葡聚糖后,可以提高斑节对虾感染白斑综合症病毒后的存活率。
本试验在饲料中添加0.2%β-葡聚糖可显著提高凡纳滨对虾血清超氧化歧化酶及酚氧化酶活力。
3.2 黄芪多糖对凡纳滨对虾的作用效果
黄芪多糖是黄芪的根系经浓缩后的干燥粉末,具有免疫促进剂的功能。研究表明,饲料中添加0.04%黄芪多糖,显著提高了齐口裂腹鱼增重率和饲料效率[6];
添加0.5%黄芪多糖,提高刺参增重率20.94%,同时也提高蛋白酶活性[27]。黄芪多糖不仅具有促生长作用,而且还影响机体的免疫机能。
在凡纳滨对虾腹部体腔注射0.1ml/尾的“鱼虾宝”(主要成分为人参皂苷和黄芪多糖)72h后,酸性磷酸酶及碱性磷酸酶活性明显上升[7]。
许杜娟[28]研究表明,黄芪多糖对脾细胞干扰素-γ和腹腔巨噬细胞肿瘤坏死因子-α的产生有显著促进作用。
本试验在饲料中添加0.05%黄芪多糖,对凡纳滨对虾增重率、消化酶及非特异性免疫因子均无显著影响,这可能与黄芪多糖的添加量不足有关。
今后应当设置黄芪多糖不同的添加量梯度,以寻求最佳添加量。
3.3 甘露寡糖对凡纳滨对虾生长、消化酶活性及血清非特异性免疫的影响
甘露寡糖是从酵母培养细胞壁中提取的一类新型抗原活性物质,是由甘露糖和葡萄糖组成的寡糖。
研究表明:饲料中添加0.2%、0.4%甘露寡糖能显著提高舌齿鲈增重率15.6%、14.5% [29];
添加0.5%、0.75%甘露寡糖提高奥尼罗非鱼增重率13.7%、13.1%,降低饲料系数0.21、0.20[10]。
在虹鳟[8]、鲤[9]、黄颡鱼[30]饲料中添加0.2%甘露寡糖均可显著提高鱼体增重率、降低饲料系数。
本试验中,饲料中添加0.4%甘露寡糖可提高凡纳滨对虾增重率12.78%(P<0.05),降低饲料系数0.09(P<0.05);
甘露寡糖对凡纳滨对虾的促生长作用,可能与改善肠道微生物菌群,促进肠道有益菌的生长,改善肠道内环境,从提高营养物质消化率有关[31-32]。
消化酶活性的增强可提高营养物质的消化率。目前,有关甘露寡糖对动物消化酶活性影响的报道较少。
在罗非鱼饲料中添加0.75%甘露寡糖,可提高干物质消化率、粗蛋白消化率14.3%、4.9%[10]。
本试验在饲料中添加0.4%甘露寡糖显著提高胃蛋白酶及肝胰脏脂肪酶活性。甘露寡糖对消化酶活性的作用,可能与甘露寡糖有益于肠黏膜上皮细胞的排列,并能改善肠绒毛高度、厚度有关[33]。
今后应当制作凡纳滨对虾消化道的组织切片,并对营养物质消化率进行测定,以寻求更直接的证据。
甘露寡糖不仅改善动物生长性能,还影响机体的免疫系统。饲料中添加0.75%甘露寡糖,可显著提高奥尼罗非鱼血清超氧化物歧化酶及碱性磷酸酶活性[10];
添加0.2%甘露寡糖显著提高虹鳟[8]、鲤鱼[9]血清溶菌酶活性;本试验在凡纳滨对虾饲料中添加0.4%甘露寡糖,可不同程度提高血清溶菌酶、酚氧化酶、超氧化物歧化酶及碱性磷酸酶活性,而这些酶是虾体重要的非特异性免疫因子。
甘露寡糖提高动物非特异性免疫功能,可能与甘露寡糖可以促进有益菌的生长、抑制有害菌的繁殖有关[31],其作用机理有待进一步研究。
3.4 蛋白酶PT对凡纳滨对虾生长和消化酶活性的影响
蛋白酶PT是一种耐高温的碱性蛋白酶,本实验室已就蛋白酶PT在水产饲料中的应用开展了较多的研究;
结果表明:在鱼粉含量为10%及6%的鲤鱼饲料中添加175mg/kg蛋白酶PT,均显著提高鲤鱼的增重率,降低饲料系数[16];在鱼粉含量为26.4%的凡纳滨对虾幼虾饲料中添加175mg/kg蛋白酶PT,可提高增重率10.8%(P=0.096) [15]。
本次试验进一步将饲料鱼粉含量降为23.0%,添加175mg/kg蛋白酶PT,结果显示提高凡纳滨对虾增重率11.25%(P<0.05),降低饲料系数0.09(P<0.05),与上述报一致。
同时本试验添加蛋白PT后,可提高肝胰脏蛋白酶、肠蛋白酶及肝胰脏脂肪酶的活性(P<0.05)。
蛋白酶PT在低鱼粉含量时改善水产动物生长性能可能是由于在低鱼粉条件下,植物蛋白含量高,可消化蛋白质降低,氨基酸组成不平衡,添加外源酶后,促进植物蛋白降解,提高蛋白质的利用率,因而提高动物生长性能及饲料利用率[16]
参考文献
[1] AI Q H, MAI K S, ZHANG L, et al. Effects of dietary β-1,3 glucan on innate immune reponse of large yellow croaker, Pseudosciaena crocea [J]. Fish and Shellfish immunology, 2007, 22: 394-402.
[2] 张辽, 温安祥. β-葡聚糖对齐口裂腹鱼生长及免疫功能的影响[J]. 动物营养学报, 2009, 21(5): 688-694.
[3] NELDA L, GERARD C, GABRIELA G, et al. Physiological, nutritional, and immunological role of dietary β 1-3 glucan and ascorbic acid 2-monophosphate in Litopenaeus vannamei juveniles [J]. Aquaculture, 2003, 224: 223-243.
[4] 许国焕, 梁友光, 吴月嫦, 等. 酵母葡聚糖对南美白对虾免疫功能的影响[J]. 饲料工业, 2003, 10(4): 53-54.
[5] CHANG C F, SU M S,CHEN H Y, et al. Dietary β-1,3-glucan effectively improves immunity and survival of Penaeus monodon challenged with white spot syndrome virus [J]. Fish and Shellfish Immunology 2003, 15: 297-310.
[6] 向枭, 陈建, 周兴华, 等. 黄芪多糖地齐口裂腹鱼生长、体组成和免疫指标的影响[J]. 水生生物学报, 2011, 35(2): 291-299.
[7] 刘宗英, 汪成竹, 覃川杰, 等. “鱼虾宝”对南美白对虾免疫相关酶的激活作用[J]. 海洋渔业, 2007, 12: 17-20.
[8] STAYK0V Y, SPRING P, DENEV S, et al. Effect of a mannan oligosaccharide on the growth performance and immune status of rainbow trout (Oncorhynchus mykiss) [J]. Aquaculture International, 2007, 15 (2): 153-161.
[9] STAYKOV Y, DENEV S, SPRING P. Influence of dietary mannan oligosacharides (Bio-Mos) on growth rate and immune function of common carp (Cyprinus carpio L) [M]//Howell B, Flos R. Lessons from the Past to Optimise the Future. Europen Aquaculture Society, 2005, 35: 431-432.
[10] 刘爱君, 冷向军, 李小勤, 等. 甘露寡糖对奥尼罗非鱼生长、肠道结构和非特异性免疫的影响[J]. 浙江大学学报:农业与生命科学版, 2009, 35(3): 329-336.
[11] 白东清, 乔秀亭. 植酸酶对鲤鱼磷钙等营养物质利用率的影响[J]. 天津农学院学报, 2003, 10(1): 6-10.
[12] 张璐, 艾庆辉, 麦康森, 等. 植酸酶和非淀粉多糖酶对鲈鱼生长和消化酶活性的影响[J]. 水生生物学报, 2009, 33(1): 82-88.
[13] 王纪亭, 万文菊, 康明江, 等. 复合酶制剂对草鱼生长性能、饲料养分消化率及免疫力的影响[J]. 大连水产学院学报, 2009, 24(5): 417-422.
[14] 钟国防, 周洪琪. 木聚糖酶和复合酶制剂PS对尼罗罗非鱼生长性能、消化率以及肌肉营养成分的影响[J]. 湛江海洋学院学报: 自然科学版, 2005, 24(4): 324-329.
[15] 刘鼎云, 冷向军, 卢永红, 李小勤. 饲料中添加蛋白酶AG对凡纳滨对虾生长和肌肉成分的影响[J]. 饲料工业, 2007, 28(20): 24-25.
[16] 冷向军, 刘鼎云, 李小勤, 卢永红,等. 饲料中添加蛋白酶AG对鲤鱼鱼种生长和蛋白质消化酶活性的影响[J]. 动物营养学报, 2008, 20(3): 268-274.
[17] LIN H Z, ZHOU J L, CHEN Y Q, et al. Effect of dietary traditional Chinese medicines on apparent digestibility coefficients of nutrients for white shrimp Litopenaeus vannamei, Boone [J]. Aquaculture, 2006, 253: 495-501.
[18] AOAC. Official Methods of Analysis. 16th ed. Association of Official analytical chemists. Washington, DC, USA. 1990.
[19] ASHIDA M, SODERHALL K. The prophenoloxidase activating system in crayfish [J].comparative Biochemistry and physiology, 1984, 77(1): 21-26.
[20] DIVAKARAN S. OSTROWSKI A. Enzymes present in pancreatic extracts of the dolphin Corphaena hippurus [J]. Journal of the World Aquaculture Society. 1990, 21: 35-39.
[21] MISRA C K, DAS B K, MUKHERJEE S C, PATTNAIK P. Effect of long term administration of dietary β-glucan on immunity, growth and survival of Labeo rohita fingerlings [J]. Aquaculture, 2006, 255 (1-4): 82-94.
[22] 陈云波, 周洪琪, 华雪铭, 等. 饲料中添加β-葡聚糖对南美白对虾的生长、存活及饲料系数的影响[J]. 淡水渔业, 2002, 32(5): 55-56.
[23] WIGGLESWORTH J M, GRIFFITH D R. Carbohydrate digestion in Penaeus monodon [J]. Marine Biology, 1994, 120: 571-578.
[24] 杨福刚, 周洪琪, 黄旭雄. 不同β-葡聚糖对凡纳滨对虾稚虾生长及非特异免疫功能的影响[J]. 上海水产大学学报, 2005, 14(3): 263-269.
[25] BAI N, ZHANG W B, MAI K S, et al. Effects of discontinuous administration of β-glucan and glycyrrhizin on the growth and immunity of white shrimp Litopenaeus vannamei [J]. Aquaculture, 2010, 306: 218-224.
[26] SUNG H H, KOU G H, SONG Y L. L.Vibriosis resistances induced by glucan treatment in tihger shrimps, Penaeus monodon [J]. Fish Pathology, 1994, 29 (1): 11-17.
[27] 孙永欣, 李亚洁, 温志新, 等. 饲喂黄芪多糖对刺参生长性能的影响[J]. 中国饲料, 2009, 4: 31-36.
[28] 许杜娟, 陈敏珠. 黄芪多糖对小鼠免疫功能的影响[J]. 安徽医药, 2003, 7(6): 418-419.
[29] TORRECILLAS S, MAKOL A, CABALLERO M J, et al.Immune stimulation and improved infection resistance in European sea bass (Dicentrarchus labrax) fed mananoligosaccharides [J]. Fish and Shellfish Immunology, 2007, 23 (5): 969-981.
[30] 于艳梅, 吴志新, 陈孝煊, 等. 魔芋甘露寡糖对黄颡鱼非特异性免疫功能及生长的影响[J]. 华中农业大学学报, 2010, 29(3): 351-355.
[31] 周红丽, 张石蕊, 贺建华. 甘露寡糖对断奶仔猪肠道菌群和抗体水平及生长性能的影响[J]. 湖南农业大学学报(自然科学版) , 2002, 28(2): 135-143.
[32] PEDROSO A A, MENTEN J F, LAMBAIS M R, et al. Intestinal bacterial community and growth performance of chickens fed diets containing antibiotics [J]. Poultry Science, 2006, 85: 747-752.
[33] 戴德渊, 黄勇富, 万红, 等. 甘露寡糖在猪生产中的应用[J] . 畜禽业: 南方养猪, 2006, 2: 24-27.
相关参考
溶菌酶抗病毒的机理是什么(2018年全国执业兽医考试真题(预防科目))
...菌体大小C、菌体排列D、细菌群体E、菌体结构答案:D102.溶菌酶杀菌作用的机制是A、裂解肽聚糖B、裂解细胞膜C、干扰蛋白质合成D、干扰核体合成E、裂解荚膜答案:A103.细菌的繁殖方式是A、芽殖B、复制C、掷孢子D、二等分分裂E...
溶菌酶抗病毒的机理是什么(2018年全国执业兽医考试真题(预防科目))
...菌体大小C、菌体排列D、细菌群体E、菌体结构答案:D102.溶菌酶杀菌作用的机制是A、裂解肽聚糖B、裂解细胞膜C、干扰蛋白质合成D、干扰核体合成E、裂解荚膜答案:A103.细菌的繁殖方式是A、芽殖B、复制C、掷孢子D、二等分分裂E...
水分测定的注意事项(阳了之后 三类食物少吃、四类食物要多吃)
...的活性,抑制其抗病能力,同时还可减少唾液,使口腔内溶菌酶减少,增加病毒和病菌在上呼吸道感染的机会。感染新冠时更要少吃盐,没有腹泻或大量出汗的一般成人每天控制在5克以内,这样有助于提高唾液中溶菌酶的含量...
溶菌酶和水解酶的区别(溶酶体为什么不会溶解消化掉自己?独特的细胞器)
...的酸性水解酶,包括蛋白酶、核酸酶、糖苷酶、磷酸酶和溶菌酶等。但是,溶酶体为什么不会溶解掉自己呢?溶酶体这个问题主要和溶酶体膜的组成有关系。溶酶体膜含有两种高度糖基化的跨膜蛋白lgpA和lgpB。这两种蛋白分布于...
溶菌酶和水解酶的区别(溶酶体为什么不会溶解消化掉自己?独特的细胞器)
...的酸性水解酶,包括蛋白酶、核酸酶、糖苷酶、磷酸酶和溶菌酶等。但是,溶酶体为什么不会溶解掉自己呢?溶酶体这个问题主要和溶酶体膜的组成有关系。溶酶体膜含有两种高度糖基化的跨膜蛋白lgpA和lgpB。这两种蛋白分布于...
水中溶解氧的测定实验报告(水中溶解氧测定意义,溶解氧测定方法-禹山传感)
溶解在水中、空气中的分子态氧称为溶解氧,通常记作DO,水中溶解氧的多少是衡量水体自净能力的一个指标。简单来说就是养殖鱼类在水体中生存呼吸所需要的氧气。禹山在线荧光法溶解氧传感器另外,对于池塘来说,提高溶...
溶菌酶溶液(2023-2029年中国溶菌酶市场研究与投资前景评估报告)
溶菌酶广泛存在于鸟类、家禽的蛋清中和哺乳动物的泪液、唾液、血浆、乳汁、胎盘以及体液、组织细胞内,其中在蛋清中含量最丰富(约0.3%)。在一些植物体如卷心菜、萝卜、无花果和微生物体内也存在溶菌酶。目前,我国溶菌...
溶菌酶溶液(2023-2029年中国溶菌酶市场研究与投资前景评估报告)
溶菌酶广泛存在于鸟类、家禽的蛋清中和哺乳动物的泪液、唾液、血浆、乳汁、胎盘以及体液、组织细胞内,其中在蛋清中含量最丰富(约0.3%)。在一些植物体如卷心菜、萝卜、无花果和微生物体内也存在溶菌酶。目前,我国溶菌...
溶菌酶是什么(2023-2029年中国溶菌酶市场研究与投资前景评估报告)
溶菌酶广泛存在于鸟类、家禽的蛋清中和哺乳动物的泪液、唾液、血浆、乳汁、胎盘以及体液、组织细胞内,其中在蛋清中含量最丰富(约0.3%)。在一些植物体如卷心菜、萝卜、无花果和微生物体内也存在溶菌酶。目前,我国溶菌...
溶菌酶是什么(2023-2029年中国溶菌酶市场研究与投资前景评估报告)
溶菌酶广泛存在于鸟类、家禽的蛋清中和哺乳动物的泪液、唾液、血浆、乳汁、胎盘以及体液、组织细胞内,其中在蛋清中含量最丰富(约0.3%)。在一些植物体如卷心菜、萝卜、无花果和微生物体内也存在溶菌酶。目前,我国溶菌...