显微镜分为哪几种(【预约试拍】超高分辨显微镜在神经科学中的应用(一))
Posted
篇首语:守望相助,一缕阳光也将照亮心扉;同心协力,一点火光也能温暖灵魂。本文由小常识网(cha138.com)小编为大家整理,主要介绍了显微镜分为哪几种(【预约试拍】超高分辨显微镜在神经科学中的应用(一))相关的知识,希望对你有一定的参考价值。
显微镜分为哪几种(【预约试拍】超高分辨显微镜在神经科学中的应用(一))
本文译自Werner等科学家在美国化学学会会刊(ACS)上发表的一篇综述,比较详实系统介绍了超高分辨率显微技术在神经科学上的最新应用进展。我们在此文基础上进行了编译整理。因文章较长,我们将分期陆续介绍。本期介绍第一部分。
《Super-resolving Microscopy in Neuroscience》
荧光成像技术在我们理解神经系统中起着关键作用。各种超分辨率显微镜方法和专用荧光探针的出现使得能够以迄今为止无与伦比的分辨率直接洞察细胞亚区室中的神经元结构和蛋白质排列。
神经元的超分辨率可视化技术揭示了对细胞骨架组成、分布、运动性和膜蛋白信号传导、突触下结构和功能以及神经元-胶质细胞相互作用的新认识。自身免疫性和神经退行性疾病模型中明确的分子靶点为使用新型和创新的成像方法深入研究疾病病理生理学提供了极好的起点。超分辨率显微镜在人脑样本和临床生物标志物检测中的应用仍处于起步阶段,但为神经学和神经科学的转化研究提供了新的机会。在这篇综述中,作者描述了超分辨显微镜在过去的二十年里是如何提高我们对神经元和大脑功能和功能障碍的理解的。

成像技术是推动生命科学几乎所有学科基础研究的核心平台。在神经科学中,共聚焦显微镜在过去的几十年中构成了用于分析神经元组织的标准荧光成像技术。固定神经元样本的激光扫描共聚焦显微镜提供了单个细胞水平上的三维和多色图像,直到树突结构的分辨率。作为补充,电子显微镜(EM)可用于获得神经元和亚室超微结构的信息,并用于脑内的连通性分析。EM非常适用于神经元突触和囊泡、细胞器和膜构象的结构分析。然而,由于靶特异性标记的方法限制,基于EM的复杂样品中的蛋白质和特异性电子密度签名的识别是有限的。作为理解神经元功能的进步,包括双光子显微镜在内的几种活体视频显微镜应用的发展使得神经元细胞培养物、器官型切片培养物的活细胞成像和动物模型的活体成像成为可能。同时,新的荧光染料、功能探针和荧光蛋白以及光遗传学方法和光学致动器(如笼状化合物)不仅允许表征,而且允许从单分子水平到整个神经系统操纵神经元及其相互作用。然而,荧光显微图像中可见细节的水平,即图像分辨率,仍然受到衍射极限的限制。
一个多世纪以来,阿贝衍射极限(定义为λ/2NA,其中λ为波长,NA为显微镜物镜的数值孔径)决定了光学显微镜的分辨率极限,并禁止分离距离小于200 nm的两个光斑。在过去二十年中,超分辨显微术(Super Resolution,SR,下文中出现SR均指超高分辨率显微成像技术)已经发展成为细胞亚衍射分辨率荧光成像和细胞器结构研究的一种非常强大的方法。SRM方法现在可以提供远低于光学显微镜衍射极限的空间分辨率,使人们能够深入了解固定的和活的神经元细胞培养制备物和组织中蛋白质的空间组织和相互作用。
本文综述了超分辨显微术的光谱学和荧光标记技术及其在神经科学中的成功应用。我们将首先提供各种SRM方法、新的荧光和功能探针以及标记技术的基本基础的详细描述。接下来作者将回顾SRM如何有助于我们理解神经元亚室结构和功能以及神经元-胶质细胞相互作用。此外,作者将概述超分辨率成像方法如何帮助我们研究自身免疫性疾病和神经退行性疾病的病理生理学。最后,作者将介绍这些新的成像方法是如何应用于分析与神经精神疾病有关的人类样本的。这些新的显微镜方法的进一步创新和日益增加的可用性将导致对神经系统功能组织的更详细的了解。

2.1.衍射屏障及其对神经科学的影响
人类大脑包含800多亿个神经元,每个神经元由数千个突触连接。因此,它构成了复杂神经元网络最突出的例子。这些网络的主要组成部分,如突触神经末梢,其空间维度接近于由约200 nm的衍射势垒所施加的分辨率限制。例如,突触活性区(释放递质的突触前细胞基质的特化区)的直径通常约为300 ± 150 nm。10突触囊泡作为运输和释放递质的关键元件,其直径甚至要小约10倍。直径为40− 50 nm。递质被释放到宽度为20−50 nm的突触间隙中,与突触后受体结合。因此,胞吐机制和跨突触信号传导的组织结构在传统光学显微镜下仍然很大程度上是隐藏的,因此需要分辨率提高约10倍(图1)。此外,位于小尺寸突触区室内的突触信号传导所涉及的大量不同分子产生了对显微镜研究具有挑战性的分子密度分布。例如,对于较小的突触,兴奋性突触可以包含几百个小泡,对于大的苔藓状纤维结,兴奋性突触可以包含多达几千个小泡,每个小泡含有多达104 - 105个递质分子。在这些小泡中,约10 ± 5个与释放位点对接,释放的递质平均遇到0−20 N-甲基-D-天冬氨酸和0−200 α-氨基-3-羟基-5-甲基-4-异恶唑丙酸受体(分别为NMDA-Rs和AMPA-Rs)在突触后侧被320 ± 130个突触后密度蛋白95(PSD-95)分子包围。

图1.衍射极限引起的光学显微镜的关键突触成分和光学分辨率极限的图示
左:兴奋性突触的中尺度绘画。突触前区室的胞质中含有大小为40 − 50 nm的突触小泡,突触前钙通道(深黄色)位于突触释放位点附近。突触小泡在突触活性区的融合是由小泡和质膜之间形成的陷阱复合物协调的(洋红色)。注意SNARE复合物在突触前活性区和插入突触前膜的钙通道附近的紧密耦合距离(青色)。突触间隙将突触前和突触后区室分开,并为跨突触相互作用的蛋白质如neurexin和neuroligin提供空间。在突触后膜中,AMPA和NMDA受体位于突触前释放位点附近。右图:左图原始图像的灰度蓝图,描绘了突触囊泡(40 − 50 nm)、AMPA-R(≈ 10 nm)、突触间隙(20 − 30 nm)的典型尺寸,以及代表光学显微镜阿贝极限的浅蓝色衍射极限斑点(≈ 200 nm,在共聚焦显微镜和低波长的最佳条件下)。
由于加速电子的波长要短得多,因此EM是唯一能够解析突触纳米级特征的方法。然而,传统EM创建了具有优异超微结构分辨率的电子密度图像,但所需的组织制备和固定以及靶向特异性标记的方法限制极大地限制了蛋白质鉴定和神经元示踪。荧光显微镜可以很容易地进行蛋白质选择性标记,但可见光(400 - 700 nm)的衍射会使所得图像模糊,并妨碍对底层纳米结构的分析。
2.2. 绕开光学衍射极限的光学显微镜方法
20世纪后期,人们开发了新的策略,通过利用物理或化学手段来区分不同荧光团的发射或减少同一时间荧光分子的数量,以尽量绕过衍射极限。减少荧光团的点扩散函数(PSF)的重叠可以通过生成光图案在集合级别以确定性方式进行,或者通过减少同一时间荧光团的数量在单分子水平上以随机方式进行。在下文中,作者将从确定性集合方法开始介绍,该方法将激光扫描共聚焦显微镜(CLSM)的有效空间分辨率推到理论极限。
2.2.1. 确定性集合超高分辨率成像方法(Deterministic Ensemble SR-Imaging Methods)
用探测器阵列代替单点探测器并打开针孔,可以将CLSM的空间分辨率提高 2倍。CLSM测量每个扫描位置探测器每个点的荧光信号。在应用适当的算法后,生成分辨率提升的图像。这些所谓的像素重分配方法包括图像扫描显微镜(ISM)、重扫描共聚焦(RSC)、光学光子重分配(OPRA)、AiryScan和即时结构照明显微镜(iSIM)。对于信号检测,使用了诸如CCD相机、光电倍增管阵列、单光子雪崩二极管阵列和六角光纤束等探测器阵列。
结构照明显微镜(SIM)在光路中插入光栅,产生相干光束与样品发生干涉,从而产生横向和轴向不同的新照明模式。然后可以使用傅里叶变换提取这种新类型的信息,从而在所有三维空间中实现空间频率解混和分辨率加倍。SIM对样品制备的要求最低,并且与所有传统的荧光探针兼容,这些探针表现出最低的光稳定性,并且可以容易地扩展到多色成像。
然而,当记录三维叠层或用于长时间成像时,强烈建议荧光团的光稳定性。此外,SIM需要的辐照强度显著较低,因此是活细胞SR实验的首选方法。为了实现更高的分辨率,非线性SIM(NL-SIM)已经引入了通过图案化饱和或荧光激发或图案化耗尽的光可切换染料。然而,对染料的开关特性的苛刻要求限制了NL-SIM对于常规生命科学实验的适用性。非线性SIM还需要更多的图像,因此实际上仅限于2D成像。另一方面,掠入射(GI)-SIM显示出高达每秒266帧的快速超分辨率成像以及100 nm的分辨率,揭示了细胞器动态的前所未有的细节。结构化照明的局限性在于其对波长的普遍依赖性、与其他SR成像技术相比的低分辨率增益,以及对系统稳定校准的需要。最后,后处理需要先验质量检查,以避免伪影,例如由于高背景信号或低效标记导致的低对比度图像而产生的人造蜂窝图案。
一种实现高得多的空间分辨率的成像方法是通过受激发射耗尽(STED)显微镜进行的超分辨率成像。在此,具有高斯分布的激发激光束被至少保留一个局部强度最小值的耗尽激光束覆盖,并用于通过扫描点外围的受激发射使荧光团返回基态。这导致了纳米级的焦点,其尺寸与耗尽光束的强度成反比,这直接转化为STED显微镜的分辨率能力,遵循:

其中λ为波长,n为折射率,α为物镜的收集角,ISTED为STED光束的辐照强度,IS为饱和强度。因此,可以通过调整耗尽激光强度来调节分辨率,在定制设计的STED显微镜中,分辨率可达到30 − 80 nm。STED显微镜模式可通过应用连续波、脉冲激发或时间门控方法来区分。采用脉冲激光的STED显微镜可降低激发能量,从而减少实时成像中的光毒性效应。通过提取隐藏在荧光团的光子到达时间中的空间信息,STED显微术中的时间选通检测也可以在较低的平均功率下操作。STED在使用商业系统时提供用户友好的高分辨率成像,无需进一步的数据后处理。实时成像(如体内脊柱动态)已得到广泛认可,但快速动态成像仍是一种点扫描方法,因此仅限于小帧尺寸。40高激光强度可能导致光损伤,但Dymin扫描和救援STED等实施方案可减少光损伤。在Dymin STED中,在共焦模式下扫描时确定最低可能的STED光束强度。根据样品的标记密度,这产生20至100倍的低STED光束强度。救援STED通过减少荧光团状态跃迁到需要分离相邻特征的区域的实施,同样减少了光漂白,因此光漂白效应减少了8倍。STED的另一个限制是其对光稳定荧光团的依赖性,因为在高激光强度下光漂白会成为问题。这对于长期动力学研究和三维叠加的获取尤其具有挑战性。值得注意的是,最近的发展通过使用荧光团标记的寡核苷酸(与连接到靶蛋白结合探针的互补寡核苷酸短暂结合)或非结合荧光团产生细胞结构的清晰负像,从而避免STED光漂白问题。两种方法,基于DNA的STED成像和超分辨阴影成像(SUSHI)分别通过荧光团标记的寡核苷酸的恒定交换和高浓度的非结合和自由扩散荧光团来防止光漂白。SUSHI已成功用于活体器官型脑切片细胞外间隙和神经纤维网结构和动力学的STED成像。
如果代替标准荧光团而使用具有毫秒寿命或更长的两个稳定状态的可逆可切换荧光团,则照明强度可显著降低。可逆可饱和/可切换光学线性荧光跃迁(RESOLFT)已通过可逆可切换荧光蛋白(reFP)实现,并成功应用于活器官型海马脑切片中树突棘的超分辨率成像。
2.2.2. 随机单分子SR成像方法(Stochastic Single-Molecule SR-Imaging Methods)
上述的确定性方法是通过改变激发模式或相位掩膜来暂时控制荧光发射达到超分辨成像,而基于单分子的定位SR显微镜则是随机地在时间上分离单个荧光团的发射。
单分子定位显微镜(SMLM)基于单个荧光团的随机激活,使用配备高灵敏相机(EMCCD或sCMOS)的宽场荧光显微镜进行单分子检测,以及精确的位置测定。通过将理想PSF与实际测量的光子分布拟合来进行分子定位。只要信号来自单个发射区,且单个发射区之间的距离大于显微镜能分辨的最小距离,则通过收集更多光子和最小化噪声,定位的标准误差可以任意小。激活和定位过程重复多次,所有定位最终用于重建超分辨率图像。
为了确保在成像的任何时候,只有稀疏的小荧光团以其活性荧光形式存在(开启状态),使用了光开关、光转换、光激活或自发闪烁的荧光团。由于定位精度和最终图像分辨率取决于每次检测到的光子数量,通常采用明亮且稳定的荧光团与>1 kW/cm2的辐照强度相结合的方式。根据所使用的荧光团不同,SMLM可达到10−50 nm横向分辨率。光激活荧光蛋白(FPs),自2006年以来已用于光激活定位显微镜(PALM),例如在405 nm的激光照射下可从关闭状态不可逆地转换为打开状态的PA-GFP和PA-mCherry; 以及可通过适当波长的激光照射从一种波长状态不可逆地转移到另一种波长状态的光转换FPs,例如MEO。此外,还成功地应用了诸如Dronpa之类的光开关FPs,其在不同激发波长的激光照射下可在非荧光和荧光状态之间可逆地切换。对于活细胞应用,使用荧光蛋白的PALM是首选方法。因为在理想情况下,每个感兴趣的蛋白质都可以用荧光蛋白进行计量标记。然而,荧光蛋白比有机染料表现出更低的光稳定性和光子计数,从而降低了定位精度,并且通常需要更长的采集时间。此外,对于PALM成像而言,融合蛋白通常会过度表达,这可能会导致不真实图像,而用转基因变体替代显示野生型表达和功能的自身蛋白仍然具有挑战性。
对于细胞内源性蛋白质的标记,通常使用有机染料的免疫标记。SMLM适用的有机染料必须是光开关、光激活或自发闪烁的,以实现单个染料发射的时间分离,但化学计量标记要困难得多。有机染料通常表现出较高的光子计数和光稳定性,从而使定位精度达到5−10nm。花菁染料Cy5和Alexa Fluor 647可以在荧光开启状态(其典型寿命为10 ms)和非荧光关闭状态(寿命为几秒,利用光开关缓冲液,缓冲液包括PBS,10−100mM硫醇,如ß-巯基乙缅(MEA),酶促氧清除剂,可以有/没有激活染料)之间可逆切换,为随机光学重建显微镜(STORM)和直接型STORM(dSTORM)的发展铺平了道路。
近年来,应用于dSTORM的染料已大大扩展,除了菁染料外,还包括罗丹明和恶嗪染料。有趣的是,最近的研究表明,即使是多个标记的抗体在光开关缓冲液中也呈现出类似于单发射的表现,因此适用于dSTORM实验。
光活化染料的作用与光活化荧光蛋白相似。也就是说,它们在被光照射或自发激活之前处于非荧光状态。罗丹明衍生物PA-JF549和PA-JF646以及桥环菁染料Cy5B是已成功用于SMLM的光活化染料。此外,在没有光开关缓冲液的水溶液中,硅罗丹明HMSiR等自发闪烁染料也能应用于SMLM。
最近,通过图案化照明方式实现更高的定位精度,单个荧光发射区的定位得到了改进。定位精度取决于信号的大小和强度,可以通过测量的PSF标准偏差的平方除以收集的光子数来估计。然而,包括拟合性能、标记密度、标记误差和显微镜漂移在内的其它参数决定了高定位精度是否可以转化为低于10 nm的空间分辨率。此外,到目前为止,因为SMLM方法成像需要昂贵的仪器和成像者具备广泛的专业知识,这在一定程度上阻碍了其广泛应用。
2.2.3. SMLM-点累计纳米成像技术(PAINT,Point Accumulation for Imaging Nanoscale Topography)
第一代SMLM技术依赖于荧光团的光开关和光激活,其分辨率需要有效地利用荧光团发出的光子数,而PAINT(point accumulation for imaging nanoscale topography)方法使用活的,与目标区域结构短瞬结合的染料。在成像过程中,被漂白的荧光团可以被成像介质中充足的新鲜荧光团不断置换替补。
由于游离染料在采集单个图像帧期间在多个像素上快速扩散,因此它们仅显示为模糊背景且不能准确定位,而结合染料显示为PSF且能准确定位。因此PAINT的第一种方法是将荧光染料(如尼罗红)与细胞膜进行非特异性结合,然后进行光漂白和新的结合。此外,基于蛋白质片段的探针被用于单分子定位标记。在最近的一个研究中,将这种方法与传统的基于phalloidin的肌动蛋白标记方法进行了比较。通过引入通用PAINT(uPAINT)使Ni-Tris-NTA与转基因蛋白质上表达的His-Tags更特异结合,并可用于突触间隙成像。uPAINT也可以应用于其它标记方法,如免疫标记(内源性蛋白抗体、纳米抗体如绿色荧光蛋白)或受体配体结合。
为了提高PAINT的适用性和特异性,引入DNA-PAINT方法。它使用长度小于10个核苷酸的短的可控的寡核苷酸链(成像链)瞬时标记其靶结合互补寡核苷酸链(对接链)。成像链与对接链的瞬时结合产生明显的闪烁。因此,荧光团开-关状态之间的切换与其光物理性质不直接关联。DNA-PAINT首先在DNA折纸(DNA-origami)上得到验证。DNA折纸是一种自组装的DNA结构(具有已知的大小),通过侧链和荧光团进行结合,并通过宽场显微镜观察。
总的来说,DNA-PAINT是一种易于实现的SR成像标记方法,无需特定光物理特性的荧光团。因为探针可以在一轮结合后,从成像介质中置换补充荧光团,从而避免了光漂白。DNA-PAINT的缺点是图像获取时间长,这是由成像链与对接链的结合和解离速率决定的,以及荧光成像链的纳摩尔浓度引起的背景信号。尽管通过使用优化的DNA序列和缓冲条件,以及使用串联的周期性DNA结构域或通过短肽的卷曲螺旋相互作用(称为“Peptide-PAINT”),可以加快采集速度,但还是要利用全内反射荧光(TIRF)(仅限于对靠近盖玻片结构进行成像的特点),才能更好地减少成像链的背景信号。
另一方面,基于DNA的探针提供了序列成像复用的明显优势,如Exchange PAINT中所述,已成功用于小鼠视网膜切片中多个结构的成像(图2)。Exchange PAINT的概念也被推广到dSTORM、STED、SIM和更传统的衍射限制的宽场和共聚焦荧光显微镜。最近,通过一种称为PRISM(probe-based imaging for sequential multiplexing)的基于DNA-PAINT的成像方法,实现了高达10个神经元蛋白质的分辨率约为20nm的多通道成像。该方法使用了低亲和力成像探针,该探针与突触、肌动蛋白和微管一抗上的对接链结合。

图2.原代神经元中几个神经元靶点的多重交换-PAINT
(A)通过DNA-PAINT显微镜连续成像的四种超分辨突触蛋白的重叠图(底行:颜色分配):圆圈表示漂移校正的基准点;不带 * 的正方形表示(B)中的高倍放大插图区域。(C)叠加(A)中标记有 * 的感兴趣区域的多路复用超分辨突触靶点和单独通道,提供关于单个靶点方向的更多详细信息。经参考文献84许可转载。版权所有2017美国化学学会。
2.2.4.定量SMLM
与所有其他超分辨成像技术相比,如果每个靶分子都可以被单独标记和定位,则SMLM还可以提供关于分子分布和分子绝对数量的单分子信息。然而,内源性蛋白质的定量免疫标记仍然是一个挑战,并且多种标记抗体的不同数量的定位可能使数据解释复杂化。另一方面,实现内源表达水平的困难以及FPs的缓慢成熟同样会使定量PALM成像恶化。然而,可以进行特别设计的对照实验以支持拷贝数估计并提取关于生物学靶结构的分子组织的真实信息。在适当算法的帮助下,SMLM可以提供关于拷贝数、聚类、共定位和复杂化学计量的数据,用于生成定量模型和模拟。此外,通过结构信息与功能突触参数的相关性也可以实现定量,例如通过生物细胞素标记膜片钳神经元。在这方面,内源性大麻素信号传导已经通过修补神经元的链霉亲和素后标记的STORM成像结合CB1受体的免疫标记,随后在GABA能海马轴突末梢内对其进行定量来研究。本研究发现,与树突投射中间神经元相比,体周投射中间神经元的CB1受体密度更高,活动区更复杂。
在黑腹果蝇中,神经肌肉接头处内源性Bruchpilot(Brp)分子的数量已经89使用抗体滴定参考实验,野生型神经肌肉接头活性区细胞基质中存在的Brp蛋白数量已被确定为约137,其中四分之三组织在约15个七聚体簇中。结合从相同组织样本记录的电生理学数据,这些数据提供了关于Brp组织如何控制活动区功能的见解。每个活动区Brp蛋白的估计数量已通过定量DNA-PAINT(qPAINT)实验得到确认,其中DNA纳米结构已用作校准参考。此外,定量dSTORM实验证明,每个活动区Brp蛋白的数量和分布受突触结合蛋白-1,的影响,突出了活动区递质释放的复杂性。在最近的一项研究中,使用Alexa Fluor 532和Alexa Fluor 647进行免疫标记的双色dSTORM已被用于量化小鼠小脑平行纤维活性区的4型代谢型谷氨酸受体(mGluR 4)的纳米级组织(图3)。本研究还使用了抗体滴定参考实验,以估计每个活性区平均含有约35个排列在小纳米域中的mGluR 4分子。此外,mGluR 4经常被发现与munc-18-1和CaV2.1通道非常接近,这支持了mGluR 4与这些蛋白质相互作用以调节突触传递的观点。这些研究显示了神经科学中定量SMLM研究的潜力,而作者没有声明其完整性。可以预见,定量SMLM的进一步发展将为突触前和突触后蛋白的组织和结构-功能关系提供有价值的信息。
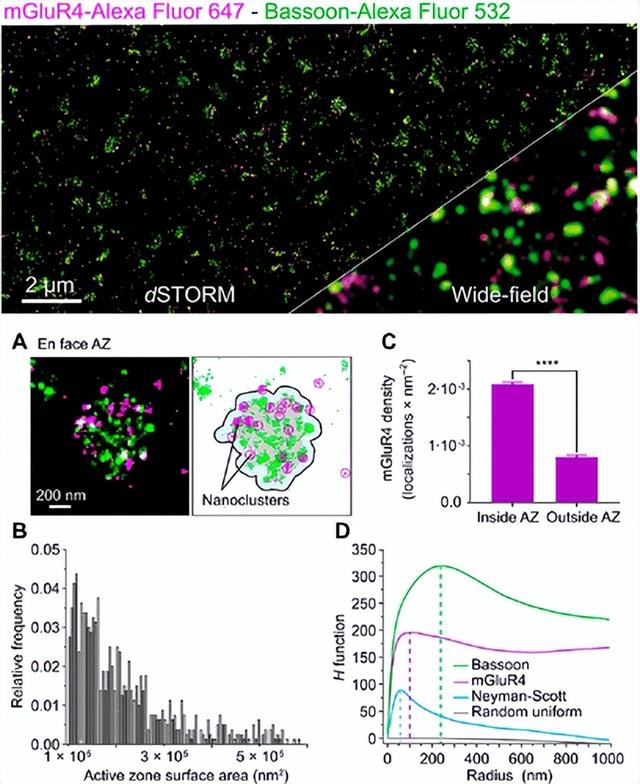
图3.在小鼠小脑切片中使用双色dSTORM记录定量代谢型mGluR4受体定位
上图:免疫染色mGluR4和巴松管的小脑冠状切片的dSTORM图像,作为活动区参考。与宽视野显微镜分辨率的比较。(A)DBSCAN聚类算法定义了正面活性区表面积(灰色)和非常接近的mGluR4信号(洋红色)。(B)活性区大小的频率分布直方图(C)mGluR4信号映射到突触和突触外区域。(D)通过Ripley's H函数分析评估巴松管和mGluR4的聚集行为。
2.2.5. 组织三维(3D)SMLM
虽然SMLM方法实现了仅几纳米的高横向定位精度,但它需要特殊的方法来打破像平面上方和下方PSF的对称性并实现高轴向定位精度。在此,PSF工程和多平面检测方案通常用于在3D中精确定位荧光团。SMLM中最常用的方法是将单个柱面透镜插入成像路径中,使PSF不对称变形,从而使用光学像散进行3D定位。基于像散的3D-dSTORM还可与光谱分层相结合,以显示COS-7细胞中网格蛋白涂层的凹坑。散光引起的畸变的程度由荧光团的轴向位置确定并因此可用于轴向位置计算。例如,3D散光SMLM已用于确定抑制性突触后密度下金卟啉和受体复合物的分布和拷贝数96或突触前活性区和突触后密度的各种成分的空间关系。双目标散光成像方案用于研究肌动蛋白、血影蛋白、和相关蛋白质的研究,发现这些蛋白质在轴突中形成周期性的环状结构,周期约为190 nm。98替代方法包括双螺旋、四足或鞍点PSF工程,使用相位掩模、变形镜、和双平面成像实现了更大的轴向范围,并已成功应用于不同的应用。− 对于在2D和3D中定位单个发射体,已经开发了不同的算法和软件工具。横向和轴向在最近的综述中列出了不同三维SMLM方法获得的分辨率以进行比较。然而,到目前为止,大多数SMLM研究都是在培养细胞上进行的,其具有相对简单的样品制备和成像要求,例如焦平面上方和下方的荧光分子的背景信号较低或没有背景信号。脑切片更具挑战性,因为它们表现出高密度的神经元结构和焦平面上下潜在的高荧光背景。由此产生的信噪比恶化是潜在地阻碍单分子定位的精确度和准确度的一个障碍。此外,脑切片必须充分透化,以实现有效的免疫标记,而不损害组织结构。除了超微结构保存外,几十微米厚的脑切片的均匀标记仍然具有挑战性。因此,最初尝试使用切成薄片的组织,这有助于标记。然而,从多个切片重建3D容积图像仍具有挑战性。为确保表位保存,在超薄切片用于SMLM成像之前,对组织进行解剖、固定以进行免疫标记、后固定、脱水并包埋在环氧树脂中。或者,3D SMLM只能对靠近盖玻片的脑切片的一个轴向平面成像。如果背景信号和散射太强,组织清除方法和基于光幕的照明方案可用于SR成像。另一个使较厚脑切片的3D成像不可靠的问题是像差。引起单个荧光团的PSF的畸变和模糊。自适应光学经常用于恢复PSF,并使各种生物体中的神经元成像更加准确。为了进一步提高基于自适应光学的SR成像的性能,将其与自适应PSF成形相结合。作为一个例子,同时校正样品诱导的畸变和产生贯穿大深度的一致PSF响应使得能够在阿尔茨海默病小鼠模型的30 μ m厚脑切片中重建纤维状淀粉样β斑块的精细细节。荧光团的3D位置还可以通过PSF内产生的自干涉来精确确定,并且已经成功地用于执行3D SMLM轴向深度达50 μ m。
参考文献(略)

目前在国内,随机光学重建显微镜STORM已成功实现商用,有需要STORM成像技术进行实验研究的专家老师们,请文末填写问卷,即可预约获得 iSTORM 超高分辨率显微成像系统试拍服务~


图、iSTORM拍摄的超高分辨率细胞图片
力显智能现已发布的超高分辨率显微成像系统 iSTORM,成功实现了光学显微镜对衍射极限的突破,使得在20纳米的分辨率尺度上从事生物大分子的单分子定位与计数、亚细胞及大分子复合物结构解析、生物大分子生物动力学等的研究成为现实,从而给生命科学、医学等领域带来重大突破。
关于我们
About us
宁波力显智能科技有限公司(INVIEW)是专业从事超高分辨率显微技术和产品研发的科技企业,依托复旦大学的自动控制、新一代信息技术及香港科技大学的生物、光学、图像处理等的技术,拥有光学、生物、自控、机械、信息技术等多领域交叉学科技术团队,将2014年诺贝尔化学奖技术产业化,推出了超高分辨率显微成像系统iSTORM、细胞智能监控助手赛乐微等一系列产品,帮助人们以前所未有的视角观察微观世界,突破极限,见所未见。
相关参考