异丙基硫代半乳糖苷(FIH催化的瞬间,受体电位锚蛋白重复结构域的羟基化)
Posted
篇首语:大道之行,天下为公。本文由小常识网(cha138.com)小编为大家整理,主要介绍了异丙基硫代半乳糖苷(FIH催化的瞬间,受体电位锚蛋白重复结构域的羟基化)相关的知识,希望对你有一定的参考价值。
异丙基硫代半乳糖苷(FIH催化的瞬间,受体电位锚蛋白重复结构域的羟基化)

文丨异文录
编辑丨异文录
图形摘要
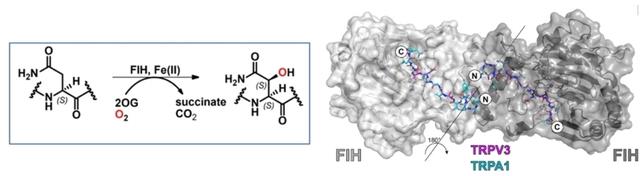
确认分析根据先前研究的不同结论,我们报道了在2-氧戊二酸加氧酶因子抑制缺氧诱导因子(FIH)的催化下,人类瞬时受体电位通道TRPA1和TRPV3锚蛋白重复结构域的天冬酰胺基羟基化的生物化学和结构研究。
摘要
瞬时受体电位(TRP)通道在动物环境感知中具有重要作用。人类TRP亚家族A成员1 (TRPA1)负责感知异硫氰酸烯丙酯(AITC)和其他亲电感觉刺激物。TRP亚家族香草素成员3 (TRPV3)参与皮肤保养。据报道,TRPV3是抑制缺氧诱导因子(FIH)的2-氧代戊二酸加氧酶因子的底物。
我们报道了关于FIH催化的TRPA1和TRPV3锚蛋白重复结构域(ARDs)的天冬酰胺基羟基化的生化和结构研究。ARD肽的结果支持了先前关于FIH催化的TRPV3羟基化的报道,并表明,在研究的12个潜在TRPA1序列中,一个序列(TRPA1残基322-348)在Asn336发生羟基化。结构研究显示TRPA1和TRPV3 ARDs与FIH结合,具有与大多数其它报道的FIH底物相似的整体几何结构。然而,TRPV3与FIH的结合方式不同于其他底物。
介绍
瞬时受体电位(TRP)蛋白是钙和钠离子通道,使细胞能够感知外部环境。TRP结构包含6个跨膜(TM)螺旋,一个位于TM螺旋5和6之间的成孔区,以及N-和C-末端胞质结构域。暴露于亲电试剂如异硫氰酸烯丙酯(AITC)会激活TRP亚家族A成员1 (TRPA1)通过修饰其N-末端胞质结构域中的半胱氨酸残基。
TRPA1的激活通常在哺乳动物中产生疼痛和刺激的感觉,而功能突变的增益TRPA1基因是家族性发作性疼痛综合征的原因。像TRPA1一样,TRP亚家族香草素成员3 (TRPV3)被外部刺激激活,包括升高的温度和化合物如香芹酚。TRPA1和TRPV3均在皮肤基底层表达,TRPV3也在皮肤健康中起作用。
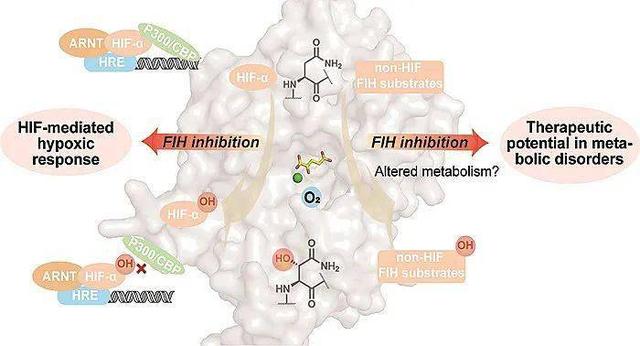
缺氧诱导因子(FIH)是一种二聚体铁二-和2-氧代戊二酸(2-OG)依赖性蛋白质羟化酶,通过催化低氧诱导因子α (HIF-α)亚单位中保守天冬酰胺残基的羟基化在低氧反应调节中发挥作用。这种翻译后修饰减少了转录活性HIF与辅激活因子组蛋白乙酰转移酶(CBP/p300)的相互作用,潜在地以基因和背景依赖的方式调节HIF转录。
也有证据表明,多个锚蛋白重复结构域(ARDs)与FIH相互作用,其中许多充当底物。值得注意的是,除天冬酰胺以外的残基(例如Asp和His)可以是ARDs中的FIH底物,而在某些情况下,FIH可以催化同一ARD的两次羟基化。
尽管HIF-α经历FIH催化的羟基化的区域在溶液中很大程度上是无序的,但是ARDs采用了良好表征的规范折叠,该折叠被提议去折叠以使得能够在FIH活性位点进行催化生产性结合。这一提议得到了生物物理分析的支持,包括FIH与HIF-α和ARD片段复合物的晶体学分析。
已经报道了TRPA1可能作为氧/缺氧传感器的证据,这是令人感兴趣的,部分是因为TRP通道在温度感应中的作用(氧溶解度随温度降低)。据报道TRPA1是HIF-α脯氨酰羟化酶(PHDs)的底物,以氧依赖方式调节HIF-α水平的催化作用。然而,对孤立博士的研究并不支持这项任务。
Karttunen等人还表明,在对分离成分和细胞的研究中,FIH可以催化TRPV3的羟基化,尽管在后一种情况下,还没有证实内源性TRPV3的羟基化。TRPV3中被提议进行FIH催化的羟基化的残基是Asn-242,其位于TRPV3胞质ARD中。重要的是,Karttunen等人提供了FIH催化的TRPV3羟基化抑制TRPV3活性的证据。
在这里,我们报道了FIH催化的TRPA1和TRPV3羟基化的生化和结构见解,其结果支持了Karttunen等人对TRPV3的研究,并表明了其他TRP通道ARDs羟基化的潜力,包括TRPA1的至少一个ARD。
实验部分
肽合成和纯化:衍生自TRPA1、TRPV3和1CA共有序列的肽使用报道的Oxyma /DIC介导的微波辅助偶联,使用CEM利伯蒂蓝固相肽合成(SPPS)机,用C-末端酰胺合成。
简而言之,使用林克酰胺MBHA树脂和0.2m FMOC保护的氨基酸的DMF溶液进行SPPS。使用20 %(v/v)在DMF中的哌嗪,随后使用CF3、三甲基硅烷、二甲氧基苯和水。使用配有Phenomenix Gemini-NX5C18 (30×250)柱的JASCO HPLC系统或配有GRACE Vydac 218TP C的Dionex Ultimate 3000 HPLC系统进行纯化18(22×250毫米)立柱。
质谱仪:MALDI-MS和MS/MS光谱是使用配备有96点研磨钢靶的AutoFlex Speed机器获得的。样品以1∶1的比例混合(v/v)比例与10毫克/毫升的α-氰基-4-羟基肉桂酸溶于50 %(v/v)含0.1 %乙腈水溶液(v/v)CF3。在装入分光计之前,让靶在室温下干燥1小时。使用Biotools 处理MS/MS数据。使用RapidFire365高通量平台和6550四极杆飞行时间质谱仪测定比活度。用Masshunter和Prism 5 处理数据。
重组蛋白生产:重组FIH以高纯度(< 95 %通过SDS-PAGE分析)生产自大肠杆菌如前所述的BL21(DE3)细胞。简言之,用编码FIH基因的pET28a(+)质粒转化感受态细胞,并在补充有卡那霉素的2-YT培养基中生长。用异丙基-β-内酰胺酶诱导表达d-硫代半乳糖苷在37℃下4小时。如所报道的,通过镍亲和和尺寸排阻色谱法纯化FIH。
射线晶体学:肽复合物结构是通过将Trp v3或trpa 1肽浸泡在预先形成的FIH⋅zinc(II)⋅NOG晶体中产生的。通过共结晶获得了fih⋅zinc(ii)⋅nog⋅trpv3(229–255)复合物结构。除非另有说明,晶体是在Intelli-Plate 96–3低轮廓平板中使用蒸汽扩散技术在20和4°C下生长的。使用Rigaku Phoenix RE滴定仪安装结晶板。
为了有氧结晶的目的,Fe2被锌取代二以避免金属氧化。fih⋅zinc(ii)⋅nog⋅trpv3(229–255)复合物的晶体通过转移到补充有20 %(v/v)甘油,并通过投入液氮中冷冻冷却。将FIH⋅zinc(II)⋅NOG复合物的晶体转移到补充有20 %(v/v甘油和Trp v3(220–246)肽或trpa 1(313–339)肽混合14小时,然后投入液氮中冷冻冷却。x射线衍射数据是在钻石光源。
使用Phaser/CCP4 (FIH PDB ID: 1H2K)通过分子置换来解析相。数据处理使用夏245并用PHENIX精制。使用PHENIX的改进和使用COOT的模型建立的改变周期被执行,直到R工作和R自由的融合了。
结果
TRPA1衍生的片段(322–348)被FIH有效地羟基化
为了探索FIH催化的TRPA1通道ARDs的范围,我们合成了一组12个推定的TRPA1衍生的含锚蛋白环的肽,用于FIH介导的羟基化分析。报道的共有锚蛋白序列(1CA)用作阳性对照(支持信息中的图S1)。之前报道的TRPV3底物也在我们的分析条件下进行了测试,发现经历了单羟基化。
这一观察结果与推定的TRPA1 HIF脯氨酰羟化酶底物缺乏活性形成对比。虽然没有明确的证据表明12种受试TRPA1 ARD肽中的11种发生了羟基化,但发现一种TRPA1衍生肽(322–348)在37°C时被FIH (2微米酶)羟基化了一次,尽管效率低于1CA,但对羟基化TRPA1衍生肽(322–348)的分析表明,羟基化发生在单个天冬酰胺残基上。
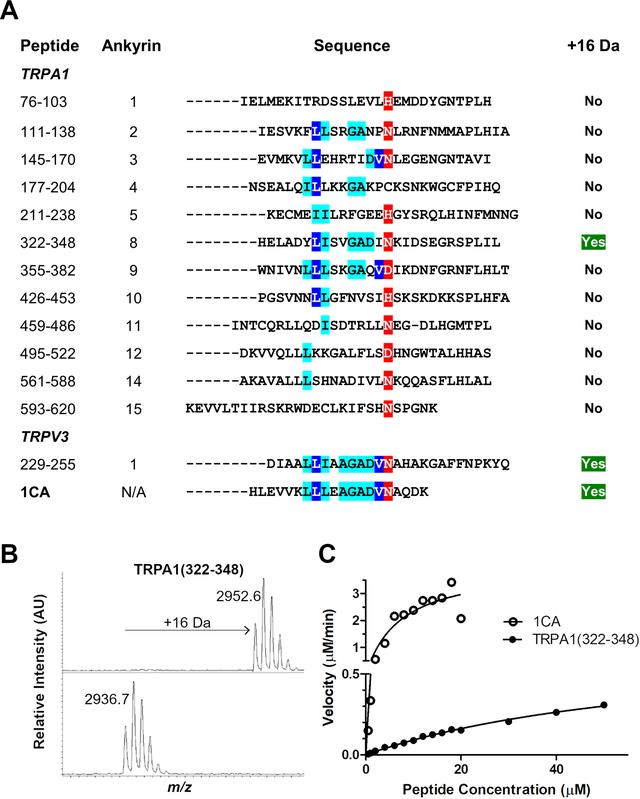
TRPA1衍生肽的FIH晶体学分析
为了研究TRP通道ARDs是否能以类似于HIF-1α和其他ARD蛋白的方式与FIH结合,我们试图在锌(II)、N-草酰甘氨酸(NOG)和Trp v3(220–246)-或trpa 1(313–339)-衍生肽的存在下结晶FIH。NOG是2-OG的近等配体,是广谱2-OG加氧酶抑制剂。
虽然在其他情况下成功,但尝试将FIH与这两种TRP通道ARD肽共结晶是不成功的。然而,在将肽浸泡到预先形成的FIH⋅zinc⋅NOG晶体中之后,在FIH二聚体的两种单体中都观察到了对应于FIH活性位点中的肽的电子密度。通过分子置换解析结构。
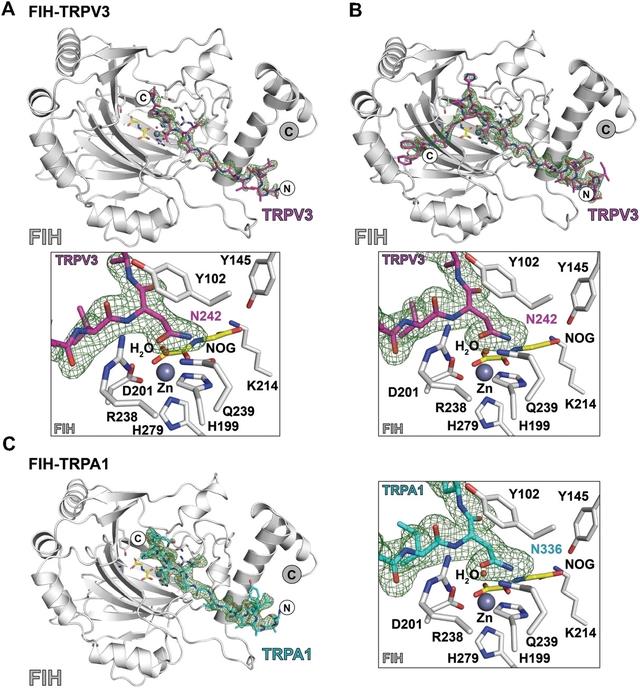
对所得结构的分析表明,TRPV3和TRPA1衍生的底物与FIH的结合方式与HIF-1αC端转录激活域的结合方式相似,这也是通过结晶学观察到的。在每种情况下,底物天冬酰胺残基通过与Gln239和Tyr102的相互作用与FIH结合,而1(相对于羟基化天冬酰胺)缬氨酸残基的异丙基被定位成以亲水方式与Trp296的吲哚环相互作用。
亮氨酸残基的侧链结合在FIH二聚化界面的疏水口袋中,如HIF-1α所观察到的。然而,只有Trp v3(220–246)肽的14个C端残基和trpa 1(313–339)肽的16个C端残基在各自的晶体结构中可见,这表明两种底物肽的N端残基在结晶状态下是无序的。二聚体中两个底物片段的可见N-末端区域残基在FIH二聚体界面区域相互投射。
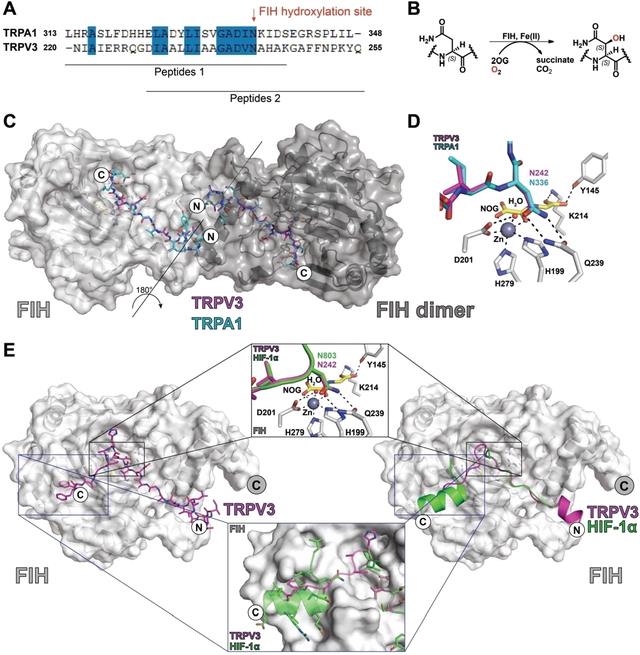
对HIF-1/2α C端转录激活域和Notch1底物的生物化学研究表明,较长的FIH底物,特别是那些在羟基化位点C端有额外残基的底物,在结构确定的底物结合位点2与FIH相互作用,可以被更有效地羟基化。
迄今为止,仅在与HIF-1α肽复合的FIH的晶体结构中观察到与FIH结合的位点2底物。为了研究TRPV3的潜在的位点2结合,FIH在Zn的存在下共结晶、NOG和在其C末端延伸的Trp v3(229-255)肽(图S3)。fih⋅zinc(ii)⋅nog⋅trpv3(229–255)复合物的最终晶体结构显示了活性位点位点1和位点2底物结合位点中底物的清晰密度。
FIH与Trp v3(220–246)和Trp v3(229–255)复合物的晶体结构比较显示,在铁螯合活性位点区域,几乎相同且明显具有生产性底物结合几何结构。仅在肽底物的末端观察到底物结合模式的灵活性,表明晶体状态下底物的活性位点结合模式基本上不受底物长度或特定ARD序列的影响。
结论
据报道,TRPA1通道活性受HIF-α脯氨酰羟化酶(PHDs或EGLNs)催化作用的调节。然而,细胞中TRPA1的脯氨酰羟基化需要进一步验证,因为随后的研究表明重组PHD1-3对包括TRPA1在内的已报道的非HIF底物缺乏活性。相比之下,我们结合的生化和结构研究支持FIH介导的TRPV3的ARD羟基化的报道,并暗示FIH可能在其他TRP通道中催化ARDs的羟基化,包括TRPA1。
FIH催化一系列ARD(和其他)蛋白质上多个残基(包括天冬酰胺、组氨酸和天冬氨酸)的羟基化。除了能够接受多种底物外,FIH还催化不同类型的氧化反应,包括去饱和和蛋白质交联。
因此值得注意的是,我们只获得了我们测试的12个TRPA1序列中的一个的羟基化的明确证据。因此,看来在FIH催化的TRP ARD羟基化中可能有高度的选择性;然而,应该注意的是,我们只研究了作为FIH底物的TRPA1的共有型序列,FIH可能与TRPA1的其他区域相互作用,并使其羟基化。
FIH催化的TRP ARD羟基化的生理学相关的生物学作用,如果有的话,目前是未知的。ARDs中天冬酰胺残基的FIH催化的羟基化可以稳定分离的蛋白质中的ARD折叠,因此,TRP通道羟基化可能调节调节蛋白的能力,例如钙调蛋白,与含ARD的TRPA1N末端胞质结构域结合。
然而,对FIH基因敲除小鼠的研究并没有揭示任何与HIF或TRP通道功能的明确联系,相反,表现出代谢率增加和体重减轻的表型。也有证据表明,FIH在HIF介导的低氧反应中的作用是环境依赖性的。因此,FIH在缺氧反应和TRP通道功能中的作用,例如在皮肤中的作用,在对FIH KO小鼠施加的测试压力下可能并不明显。然而,由于缺乏FIH催化的其他ARD蛋白羟基化的明确功能分配,这种可能性可能被认为是推测性的。
对FIH与TRPV3和TRPA1 ARD片段复合物的结晶学研究显示,结合方式与HIF-1α和先前研究的ARD FIH底物的结合方式相关,尽管它们提示TRPV3与FIH的结合方式可能与远离直接活性位点区域的其他底物的结合方式不同。
观察到一个延伸的TRPV3片段与FIH的底物结合位点2结合,其结合方式与HIF-1α C端转录激活结构域的结合方式有些不同。位点2的结合可能有助于调节不同底物被FIH羟基化的速率。迄今为止,尚未报道TRPA1的锚蛋白-8的结构;然而,TRPV3的结构分析表明,TRP通道(和其他ARD FIH底物)的ARDs需要,至少部分地,去折叠,以便有效地与FIH结合,正如与FIH结合的其他ARD蛋白的催化生产性结合的情况。
TRPA1和TRPV3组装成同源或异源四聚体以形成功能性通道,一种可能使几个TRPA1和TRPV3亚单位的ARDs接近的排列。FIH在溶液中形成同源二聚体,该同源二聚体至少在晶体状态下可以同时结合两种底物。
尽管在我们的晶体结构中没有观察到同时与FIH二聚体复合的两种TRPV3/TRPA1底物之间的直接相互作用,两个结合底物的N-末端区域非常接近(> 7)。因此,FIH二聚体有可能同时与来自相同或不同TRP通道的两种ARDs相互作用。
笔者认为:鉴于高等动物中高度保守的FIH的生物化学和多重底物作用的明显复杂性,我们认为调节FIH活性的治疗潜力,包括TRP通道活性,最好通过开发与疾病相关的体内试验相关的有效小分子抑制剂来探索。
参考文献:

相关参考